
I HAVE BEEN planning on writing this post for quite a while and hope to do the topic justice. The Cambrian explosion refers to the sudden radiation of metazoan life during the Cambrian period, and is usually placed between about 540-530 million years ago. It is often said that during the Cambrian explosion all of the modern phyla appeared. This statement has been seized upon by creationists who claim that the Cambrian explosion proves a sudden supernatural creation of life, rather than evolution of organisms over time.
No doubt you have noticed that this blog is pro-evolutionary, but I have not spent much time in the past being anti-creationist. I do not expect to devote much time to the topic of creationism in future. This isn’t because of disinterest, because as an ex-creationist I probably pay more attention to the topic than most scientists. It is more that I think there is so much to be said for evolutionary science and consider it more profitable to devote time to real science here.
You’ll note that creationists do not take the same tack. If creationism had any scientific merit whatsoever you would find creation scientists hiking around in the Karoo digging up basal synapsids, explaining what their ecosystem was like, and explaining how this Permian ecosystem could remain so segregated from the many supposedly contemporary Triassic, Jurassic, Cretaceous, and Tertiary ecosystems such that these plants and animals never invaded each other’s areas, and how in the world they managed to sort themselves so neatly during the Flood. In fact there is a glaring lack of such activities, and creation “science” largely proceeds by trying to poke holes in evolutionary theory. Thus they seize upon the Cambrian explosion. “The fossil record shows the sudden appearance of the phyla, so evolution must be wrong!” they say. This is perplexing in its own way, since creationists spend most of their time denying the validity of the fossil record, and attempting to criticize evolution on the basis of the Cambrian explosion requires acceptance of its validity. But when you are arguing against one model rather than in favor of an alternate model, consistency inevitably suffers.
So what do we actual see in the Cambrian explosion? Did all modern phyla appear there? Is this a killing blow for the allegedly perpetually tottering, yet somehow never toppling giant of evolutionary theory?
Once again, you’re probably not all on tenterhooks.
One thing that I’ve been thinking about more lately is placing extinct species in context in their ecosystems. I’ve tried to do that in recent posts examining what saber-toothed cats ate and why Simocyon retains tree-climbing ability. This is difficult because it requires a knowledge of that species’ contemporaries, both animals and plants (I admit to be hopelessly biased in favor of animals). It turns out that the Cambrian ecosystem was much different than any ecosystem today, and would be unable to persist in the presence of competition by modern organisms if we could recreate a Cambrian ecosystem in modern times.
During the Cambrian multicellular life only existed in the sea. This is hard to imagine. The continents were bleak–sand and rock, with the only living organisms being bacterial crusts.1 In the oceans plant life was represented only by algae, mostly single-celled and filamentous, but with some small branching forms. But the key feature of the Cambrian were thick bacterial mats that covered shallow seabeds. Such mats only grow in unwelcoming habitats now. At that time the mats prevented vertical tunneling into the substrate. This type of tunneling is ubiquitous in most modern environments, and this bioturbation produces characteristic patterns in layers of strata. Bioturbation did not become common until late in the Cambrian, when organisms had developed adaptations allowing tunneling into this dense and oxygen-poor substrate.2 Many Cambrian organisms crawled about grazing upon this microbial mat.
The organisms of the Cambrian were also unusual. Some elude classification, others have at times been placed dogmatically in one group and then later placed just as dogmatically in a different group. This is due to a flaw of Linnean taxonomy. It is an either-or system that works well for distinguishing modern organisms, but is not designed to communicate evolutionary relationships, and can even help in obscuring them. This is not to say that it is not useful, we just need to remember that there are certain things that Linnean taxonomy does not tell us.
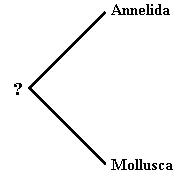 We know that the bilateria are monophyletic, so all phyla must come from a common ancestor. Let us examine a cladogram showing annelids at one tip and molluscs at the other. Modern examples of these organisms are easily distinguished. But if we trace down the branches to the root of the tree, the two phyla share a common ancestor. If we were to examine this ancestor, would it be an annelid or a mollusc? It would not really be either. We would expect it to have traits of both, but perhaps also to lack key traits, or include additional traits. If a person discovered a fossil of that common ancestor and was asked what phylum it fit into, that person would probably place it in either Annelida or Mollusca, depending on which traits were given priority. Likewise, an organism slightly up the branch towards modern molluscs might retain enough of the traits of its common ancestor that it might also be placed in Annelida, or vice versa. If these transitional species were not long extinct, we probably would not have two phyla at all, but one phylum consuming Annelida, Mollusca, and all of the ancestors of both down to their common ancestor.
We know that the bilateria are monophyletic, so all phyla must come from a common ancestor. Let us examine a cladogram showing annelids at one tip and molluscs at the other. Modern examples of these organisms are easily distinguished. But if we trace down the branches to the root of the tree, the two phyla share a common ancestor. If we were to examine this ancestor, would it be an annelid or a mollusc? It would not really be either. We would expect it to have traits of both, but perhaps also to lack key traits, or include additional traits. If a person discovered a fossil of that common ancestor and was asked what phylum it fit into, that person would probably place it in either Annelida or Mollusca, depending on which traits were given priority. Likewise, an organism slightly up the branch towards modern molluscs might retain enough of the traits of its common ancestor that it might also be placed in Annelida, or vice versa. If these transitional species were not long extinct, we probably would not have two phyla at all, but one phylum consuming Annelida, Mollusca, and all of the ancestors of both down to their common ancestor.
Referring back to my post on the article “Understanding Evolutionary Trees”, two common mistakes in analyzing evolutionary trees are assuming no change along a branch and assuming change only at nodes. Both of these mistakes contribute to misunderstandings about the Cambrian explosion. Budd and Jensen point out the problem of assigning long-extinct groups to existing phyla.3
Extant monophyletic groupings are always morphologically distinct from their extant sister-group, and that distinctness is brought about by subsequent extinction of the lineages (plus its offshoots) that led to each of them, away from their last common ancestor. As random extinctions through time slowly remove lineages, the most basal taxon of a clade will sometimes be the victim, thus widening the path-length between the surviving most basal members of extant sister clades (Fig. 3). The bases of clades are therefore eroded by extinction, and, as only living members of the clade can rediversify, this is a permanent loss. These extinct basal taxa will not possess all of the apomorphies that define the basal node of the surviving clade. It should be noted that this process will occur whether or not basal members of clades are particularly prone to extinction or not; there does not have to be anything ‘‘special’’ about basal taxa. One further aspect about these now extinct basal taxa is that they would have accumulated their own autapomorphies not possessed by the extant taxa. As a result, these basal fossil taxa are bound to differ from the extant clades: they will not be diagnosable as members of those clades; and they will show a confusing mixture of some but not all features of those clades, together with a set of features absent from them. It should be noted that this characteristic mix has been repeatedly noted in Cambrian fossils. For example, Hughes (1975) said of the Cambrian arthropod Burgessia: ‘‘what is apparent from this restudy is that Burgessia did possess a mixture of characters . . . many of which are to be found in modern arthropods of various groups’’ (Hughes, 1975, p. 434).
This diagram3 shows how this erosion takes place.
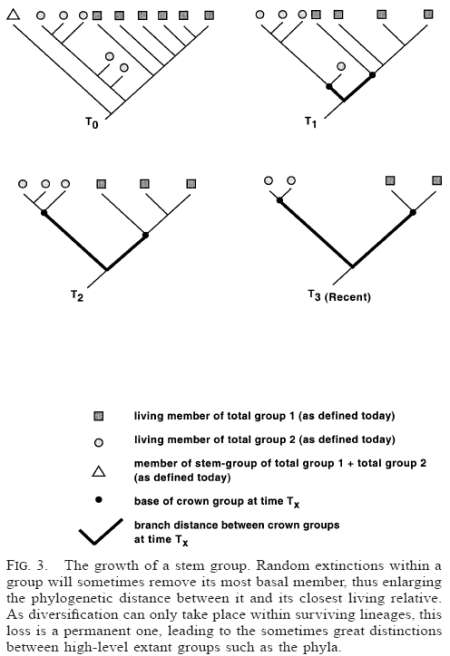
 The way to help solve this problem is to recognize there is a distinction between crown groups and stem groups. Crown groups contain extant organisms, while stem groups are made up of extinct groups that may lack some crown group features, but also may retain features that were present in the common ancestor and later lost. As you can see from this diagram, if we extrapolate from our knowledge of extant species to determine what traits are diagnostic for their clade, we may unintentionally slip into the assumption that no significant change occurred since their divergence, and pick a trait to be diagnostic of the clade that in reality was not characteristic of the basal members of that clade.4 This can lead to exclusion of a group from its proper phylum, inclusion of a stem group more closely related to a different phylum in the wrong phylum, or the generation of a redundant new phylum to contain a problematic group.
The way to help solve this problem is to recognize there is a distinction between crown groups and stem groups. Crown groups contain extant organisms, while stem groups are made up of extinct groups that may lack some crown group features, but also may retain features that were present in the common ancestor and later lost. As you can see from this diagram, if we extrapolate from our knowledge of extant species to determine what traits are diagnostic for their clade, we may unintentionally slip into the assumption that no significant change occurred since their divergence, and pick a trait to be diagnostic of the clade that in reality was not characteristic of the basal members of that clade.4 This can lead to exclusion of a group from its proper phylum, inclusion of a stem group more closely related to a different phylum in the wrong phylum, or the generation of a redundant new phylum to contain a problematic group.
Upon further examination, most Cambrian organisms either have a mix of traits or have some traits diagnostic of a crown group, but lack other crown group traits. These organisms are more accurately placed as stem groups of the modern crown group phyla. Budd and Jensen point out that for most phyla crown group members do not begin to appear until 20-25 million years after the beginning of the Cambrian explosion, well after the “explosion” was supposedly finished. The crown groups evolved through the gradual acquisition of some traits and loss of others over tens of millions of years. This is quite a different picture from the creationist representation of the Cambrian explosion as an extremely rapid emergence of distinct modern phyla, with nary a transitional to be seen.
Indeed, most of the organisms we consider most notable did not appear in the Cambrian. Plants are the most obvious example to me–the Cambrian landscape was barren, if we overlook microscopic bacteria. Land plants are now key producers and have a major impact on local ecology. Most plant divisions (the equivalent of phyla) evolved after the Cambrian. Plants first began to colonize the land during the Ordovician following the Cambrian, but the vascular plants did not evolve until the Silurian, 425 million years ago. Through the Carboniferous (360-300 million years ago) tons of coal were laid down from giant fern forests, although the first gymnosperms also began to appear. These plants dominated through the Permian and Triassic, but in the Jurassic (200-145 million years ago) began to be replaced by angiosperms. The angiosperms, flowering plants, are the major group of plants that we observe, including almost all deciduous trees (with exceptions such as the larch, a deciduous gymnosperm), bushes, and herbs. The grasses are angiosperms and appeared during the Cretaceous (145-65 million years ago), but did not become ubiquitous until the middle of the Tertiary, only 10-20 million years ago. And much more could be said for the evolution of vertebrates, including the extremely diverse fish and the myriad land-dwelling tetrapods. Amphibians, reptiles, mammals, birds, and all of their extinct tetrapod ancestors appeared long after the Cambrian. Even some organisms we consider “primitive”, such as insects and spiders, appeared about a hundred million years after the Cambrian explosion. The Cambrian may have been the time period that saw key metazoan phyla diverge, but it was by no means the beginning of modern life!
Not only did all modern crown groups not appear in the Cambrian explosion, but the Cambrian explosion was also not the appearance of metazoan life. The Cambrian was preceded by the Ediacaran (formerly the Vendian), and we have fossils of organisms from the Ediacaran that are even stranger than those appearing in the Cambrian. We have trouble placing Cambrian organisms in modern phyla, but we sometimes have trouble placing Ediacaran fossils in the right kingdom. Considering what we know about the divergence of the phyla in the Cambrian, this is expected. The Ediacaran is closer to the convergence of the kingdoms, and contains organisms long extinct that possess characteristics not possessed by organisms that evolved later. Some scientists have proposed that most of the Ediacaran fauna be placed in their own phylum since it is so hard to diagnose their affinity to modern organisms.
While the Ediacaran biota shared a common ancestor with all of life, we may never be able to determine their exact place on the tree of life. The fossil record for the Ediacaran and Cambrian is spotty, with only a few places preserving detail of soft-bodied organisms. It is possible with the discovery of new fossil-bearing formations that we will be able to determine the evolutionary relationship of the Ediacaran biota to the Cambrian biota, and thus to all metazoan life. Even if we are not able to interpret the Ediacaran as clearly as we would like, we will make progress with the Cambrian biota. The recent discovery of several formations with excellent morphological preservation of Cambrian organisms will contribute to the study of the evolution of the phyla, as we place stem group members and perhaps eventually converge on the common ancestors of modern phyla.
- Labandeira, C. C. “Invasion of the continents: cyanobacterial crusts to tree-inhabiting arthropods.” Trends in Ecology and Evolution 2005, 20, 253-262. DOI:10.1016/j.tree.2005.03.002
- Dornbos, S. Q. “Evolutionary palaeoecology of early epifaunal echinoderms: Response to increasing bioturbation levels during the Cambrian radiation.” Palaeogeography, Palaeoclimatology, Palaeoecology 2006, 237, 225-239. DOI:10.1016/j.palaeo.2005.11.021
- Budd, G. E. “The Cambrian Fossil Record and the Origin of the Phyla.” Integrated and Comparative Biology 2003, 43, 157-165. DOI:10.1093/icb/43.1.157
- Budd, G. E.; Jensen, S. “A critical reappraisal of the fossil record of the bilaterian phyla.” Biological Reviews 2000, 75, 253-295. DOI:10.1017/S000632310000548X

8 comments
Comments feed for this article
March 21, 2008 at 11:43 am
Don
You’re arguing against the notion that modern phyla started with the Cambrian explosion, but not against the notion of the explosion itself. Correct?
Your point about the problems with shoe-horning fossil species into modern categories is a good one. I guess this bias of the classification scheme that Linnaeus came up with makes sense because he developed it at a time when species were thought to be static and eternal things.
March 21, 2008 at 5:00 pm
Christopher Taylor
To be honest, though, I’ve never quite been able to decide to what extent Budd & Jensen had a valid point, and to what extent they’re just waving the magic re-definition wand to make the problem go away. There was a significant change in ocean ecology and the taxa involved in it during the Cambrian, but the best way to describe that change is a bit more difficult…
It’s a little unclear what Linnaeus himself thought on this subject. He certainly didn’t accept common descent for all organisms as a whole, but he may have accepted the possibility of speciation below about the ‘genus’ level.
The ‘shoe-horning’ problem of Linnean taxonomy is a side-effect of the insistence that all taxa must be assigned to certain ranks. So a common ancestor of annelids and molluscs still must belong to a phylum – either one of the daughter phyla, or an extinct phylum of its own. None of these options are particularly good at indicating evolutionary relationships.
March 21, 2008 at 5:30 pm
Nimravid
The Cambrian explosion was a real radiation of pre-existing life forms, but the organisms produced during the explosion then evolved gradually to eventually gain all of the characteristics possessed by modern members of their phylum. By the end of the Cambrian explosion most phyla still had some millions of years of evolving to do before the modern body plans were reached.
Which is fine. I’ve read about some of the plans to try to replace Linnean taxonomy with some method combining phylogenetics, and it could get really messy. As long as we know what the goal of taxonomy is it should not be misleading.
March 24, 2008 at 11:14 am
Stefan Patejak
Even if the Cambrian explosion does refute evolution, it still doesn’t fit with creationism. What it would say is that starting about 530 million years ago, the Flying Spaghetti Monster created various lifeforms over a period of 70 million years. (Presumably, on the 70,000,001st year he rested.) Furthermore, these lifeforms do not include most of the ones we see today. It makes no sense that a sentient being would do this, and it certainly doesn’t fit with Genesis.
June 5, 2009 at 8:48 pm
Gary Navarre
The Cambrian Explosion fits well enough with Genesis if you get rid of the fanciful “530 million years ago” traditional geological assumption (dreamed up primarily to accomodate Darwinism and Uniformitarianism), and replace it with a “10,000 years ago” assumption (followed in turn by a “Great Flood” assumption).
Of course this is almost as radical as the “flying spaghetti monster” but both seem preferable to trying to keep alive the myth that natural selection, mutation, et al., can adequately account for the speciation we see today. It now seems clear beyond reasonable doubt that natural selection (et al) is unable to increase the amount of genetic information available to a species, and therefore tends to promote stasis in a species rather than “evolution.” Thus, if we are to be truly objective, the available evidence suggests that the Cambrian phyla were indeed “planted” on earth and already carried all the genetic information necessary for micro-evolution and adaptation to any variety of earthly habitats. But favored paradigms die hard, no matter how forthright the evidence against them.
July 24, 2009 at 11:59 am
Texoki
¿Can you name the transitional fossils ancestrors of the 50 Phylum of the Cambrian Explosion?
December 27, 2009 at 2:16 am
Gary Navarre
The rocks aren’t cooperating with the evolutionists either. Consider a recent finding regarding the origins of granite, which has long been a problem for geologists. The current evolution-based theory (invented out of thin air), states that the granite structure of the earth’s crust formed as magma cooled over hundreds of millions of years. This assumption is fundamental to current theories of earth’s vast age.
Along comes Polonium 218, a radioactive isotope found in granite all over the world. As it decays it creates a “halo” which is visibly preserved in granitic rock and can be viewed under the microscope (pics available on net). These halos are the actual physical record of the emission of atomic particles as the polonium nucleus decays. Note that these halos are immediately erased by high temperatures (over 150 degrees C.). When granite is melted in the lab the halos are erased totally and the granite cools into rhyolite – not granite. Thus we can be sure that the halos were present primordially – at the original formation of the granite.
A creation scientist discovered that the half-lives of the Polonium isotopes are only 3 minutes (+ ½ = 3 min for 218 Po), which is too short to allow for anything but an extremely rapid mineral crystallization. Anything over 3 minutes would erase the halo. To make a long story short, their presence demonstrates that all the granite in the earth’s crust was formed in less than 3 minutes.
Yes- you heard me right. So objectionable was this discovery to the evolutionist establishment that the National Academy of Sciences tried to bury it. But finally, after full and repeated peer review it has been published in some of the most prestigious scientific journals. Listen to some of their shocking (but welcome) admissions:
“Current physical laws may not have governed the past “
“Early primordial crustal rocks, rather than cooling and solidifying over millions of years, crystallized almost instantaneously.”
“Some geological formations thought to be 100 million years old are in reality only several thousand years old.”
“Grant these propositions – and any researcher will tell you the entire structure of the historical sciences would dissolve into themselves. Few certainties would remain. Yet these possibilities and others equally disintegrative have been published over the past several years in the world’s foremost scientific journals [including] Nature, Science, the Annual Review of Nuclear Science, among others.”
We also find confirmation of these experimental results in uranium halos in granite (for different reasons) and through mass analysis of zircons extracted from granitic cores. Through this discovery and others, evolutionary theories of earth’s formation are out the door – along with that of the whole solar system Oh well. Evolution contradicts every principle of science from the 2nd Law of thermodynamics to the simple laws of statistical probability, so perhaps its time for a revolution. Suppose those cambrian rocks are only about say, 6000 years old? The howls of protest from the establishment continue but to date there has not been a single peer-reviewed publication to refute these experimental findings.
The coyotes howl but the wagon train rolls on. Try this theory of earth;s past: For he spake, and it was done; he commanded, and it stood fast.
(Psalm 33:6,9)
September 23, 2010 at 4:19 pm
Texoki
Can anybody name the transitional fossils ancestors
of the 100 Phyla of the Cambrian Explosion?